Translate this page into:
Attentional modulation of heart rate variability (HRV) during execution of PC based cognitive tasks
Abstract
Spectral analysis of HRV was contemplated in 15 young male subjects during their engagement in PC based Cognitive Tasks [consisting of Auditory Tone Discrimination, Working Memory and Continuous Performance (Vigilance) Tasks, each with two or more levels of memory load/time compression]. Manipulation of cognitive demands in these task variants was objectively defined. Results of the study did establish susceptibility of certain spectral components of HRV to cognition especially when locus of the load was on working memory. The effect was demonstrably independent of possible entrainment of power spectrum of HRV with the stimulus frequency and/or changes in breathing pattern. Nevertheless, a change was demonstrable only between ‘rest’ and ‘task’ conditions and it did not co-vary with a further manipulation of load in the working memory. Modulation of HRV during cognition seemed to be correlational and not causational. Results are discussed in the text.
Keywords
HRV
Cognitive tasks
Attention modulation
Auditory tone discrimination tasks

Several studies, under laboratory conditions,have shown that increasing mental load and attention cause a decrease in both time and frequency domain estimates of heart rate variability (HRV) [1,2,3,4,5]. A decrease in HRV during attentional phases (and its return to pre-stimulus level approximately five seconds following attention termination) is reported even in young infants [6]. O Hanlon [7] has confirmed these findings in a passive monitoring task and, moreover, he found that when fatigue reduced subjects attention, the initial decrease was changed into a gradual increase in HRV. Nevertheless, there are observations wherein HRV was found not to be susceptible to mental workload demands [8,9,10] especially in a realistic environment.
The present study was conducted with an aim to examine whether a demonstrable change occurs in HRV (and its constituent spectra) consequent upon engagement of subjects in tasks which can be typified as mentally loading and whether it co-varies with a further manipulation in cognitive demands of the tasks.
Standard physiologic texts recognise three main components in a HRV spectrum calculated from Short recordings of 2 to 5 minutes. These are very low frequency (VLF) (upto 0.04 Hz), low frequency (LF) (0.04 - 0.15 Hz) and high frequency (HF) (0.15-0.40 Hz) components [11]. The physiologic explanation of VLF component is much less defined and the existence of a specific physiologic process attributed to these heart period changes might even be questioned. The non-harmonic component which does not have coherent properties and is affected by algorithms of baseline or trend removal, is commonly accepted as a major constituent of VLF. Thus VLF assessed from short-term recordings is a dubious measure and is recommended to be avoided when HRV of short term ECG is interpreted [11]. The HF component reflects momentary respiratory influences on the heart rate, is decreased by tilting or parasympathetic blocking drugs and is increased by sympathetic blocking drugs or controlled respiration. Therefore, the HF component in humans has been thought to provide a quantitative and specific index of vagal modulation [11,12, 13]. Interpretation of LF components is, however, controversial. The LF component is increased while standing and the increase is blocked by intravenous propranolol [12,13]. Moreover, this component is not found in quadriplegic humans who have severe dysfunction of the sympathetic nervous system [14]. Therefore, the LF component has been interpreted as an indicator mainly of sympathetic influences (especially when expressed in normalised units). Consequently, the LF/HF ratio has been reported to be a convenient index of sympatho-vagal interaction [11]. Nevertheless, it is also considered as a parameter that includes both sympathetic and parasympathetic influences [15].
All the components have been reported to decrease under conditions of increased mental activity.
Methodology
15 young, male, nonsmokers, technical graduates were evaluated during their engagement in PC based Auditory Tone Discrimination Tasks (two Variants), Working Memory Tasks (three variants), and Continuous Performance (Vigilance) Tasks (two variants). The subjects were ascertained to be healthy by history, a thorough clinical examination and resting ECG. In the above tasks, memory load and temporal demands were manipulated. The subjects were explained regarding the experimentation and its relevance and an informed consent was obtained. They were asked to observe total abstinence from alcohol since previous night (from the day of experimentation) and have a sound sleep. Physical attributes of the subjects were as follows - Age - 24 ± 2 years, Height - 17 ± 7 cm and 59 ± 7 kg. (Values are mean ± SD.
In Auditory Tone Discrimination Tasks, a randomised series of two tones was presented through a computer which was kept at a distance of 50 cm from the subject anteriorly in the midline. The tones varied in frequency and probability as follows- 1,000 Hz [0.67] (standards) and 2,000 Hz. [0.33] deviants). The tones were 50 ms bursts presented at 40 dB. Subject was to discriminate between the two tones and respond to the occurrence of the deviants, by a key press. Two such tasks were administered. Time compression in the Task-2 was manipulated to be 4 times more than that in the Task -1 by decreasing the inter-stimulus delay from 2000 ms to 500 ms in Task 1. Performance was evaluated by comparing the number of deviants reported by the subject against the figures provided by the Computer Programme at the end of experiment. In Working Memory Task, the subject was to identify a target stimulus in an array of similar stimuli. The stimuli was one of the 90 geometrical figures of different shapes and of the size of 3.8 x 2.5 cm. The arrays comprised of 4 such stimuli (two on the left and the other two on the right of the array) arranged horizontally with an inter-stimulus separation of 0.6 cm. One of these stimuli could be the target stimulus. The subject was asked to indicate, by different key presses, if the target appeared in the right or the left of the array or it did not appear in the array at all. All these occurrences were kept equiprobable in the task programme. There were two unique features of the task; first, the subject had to wait till the disappearance of both target and array before executing a response (else the response was not accepted by the Programme) and, secondly, the next target appeared only when the subject had responded. Pre-target delay, pre-array delay, inter-stimulus delay and exposure time of the target were varied to manipulate the time compression and/or memory loads. The above characteristics of the three variants of the Working Memory Task are given below :-
| Task-1(WM-1) | Task-2(WM-2) | Task-3(WM-3) | |
|---|---|---|---|
| Pre-target delay (ms) | 500 | 500 | 200 |
| Exposure tune of target (ms) | 500 | 200 | 500 |
| Pre-array delay (ms) | 500 | 500 | 500 |
| Exposure time of array (ms) | 1000 | 400 | 1000 |
| Intrer - stimulus delay (ms) | 500 | 500 | 200 |
In Continous Performance (Vigilance Tasks), subject was to recognise a consecutively repeated appearance of any target and respond by a single key press. The target stimuli were from a group of 90 geometrical figures of different shapes and of the size of 3.8 x 2.5 cm. Exposure time of target in the two variants of the task was 400 and 200 ms. The corresponding values of inter-stimulus delay were 1000 and 500 ms.
Duration of each task variant was 5 minutes.
ECG data in standard Lead-II configuration were acquired, continuously, using impedance cardiograph (ICG-M501 ASK Ltd, Budapest, Hungary) from Scalene Engineers Pvt Ltd., Bangalore - 560 071. ECG signal was band passed through a filter (0.05-50 Hz), digitised with a sampling frequency of 500 (± 0.1%) Hz with 12 bit resolution and stored on the hard disk of an IBM -compatible 486 DX2 Personal Computer for off line analysis. Detection of R wave peaks was through IKX V5.29 software (ASK Ltd. Budapest, Hungary) using ICG-M501 data. Sequence of beat to beat RR intervals was manually examined and edited for artifacts or ectopics following which this discrete even series (DES), was transformed into an evenly sampled time signal at 4 samples/sec by linear interpolation using Matlab* (Version 5.1.0.421) software. An estimate of HRV was made in Matlab* (Version 5.1.0.421) software with ‘SPECTRUM’ command using Welch’s averaged periodogram method wherein the length of signal was divided into overlapping sections, each of which was detrended. Length of section was 256 with 50% overlap . A Hanning window was used. Discrete Fourier transforms of the sections were averaged and the out put was in the form of spectral densities. Low and high frequency power were quantified by integrating the spectra as follows - VLF - up to 0.04 Hz; (i.e., Total power in the spectrum minus that in the VLF component). VLF power was not processed/analysed further due to its dubious (and even questionable) association with any physiological processes [11]. Additionally power was also calculated for mid frequency band (0.07-0.14 Hz) as it has also been reported to be useful in psychophysiological texts [16]. The LF and HF power were expressed in absolute terms (ms2) and in units normalised relative to total power (minus the VLF component) [11] as well as squared mean RR interval [17]. LF/HF Ratio was also computed.
Statistical analysis
To ascertain the significance of difference in performance (accuracy and response latency) between the two Auditory Tone discrimination and Continuous (Performance Vigilance) Tasks, paired ‘t’ test was used. Univariate Anova with repeated measures was used to see the differences in performance amongst the three Working Memory Tasks. Following a significant out come from ANOVA, post-hoc analysis was done using Tukey’s HSD. Wilcoxon Matched Pairs test was used in the rest of the analysis wherein the data distributions were not normal and application of a parametric static was not applicable. This decision was made subsequent to examination of distribution using Shapiro Wilk’s ‘W’ Statistic. All values are presented as Mean ± SD.
Results
Performance of subjects is given in Table-1. Results of power spectral analysis of HRV are given in Fig-1 and Tables-2 to 4.
| Task-1 | Task-2 | Task-3 | |
|---|---|---|---|
| Auditory* | 98.7 ± 2.0 | 99.2 ± 1.0 | - |
| Working Memory** | 92.7 ± 5.2 | 84.5 ± 5.5 | 93.4 ± 5.2 |
| Vigilance* | 82.1 ± 12.9 | 81.7 ± 14.8 |
p-0.000, 0.921 and 0.000 for Working Memory Task-1 vs 2, 1 vs 3 and 2 vs 3, respectively (Post-hoe Comparisons using Tukey’s HSD)
| Task-1 | Task-2 | Task-3 | |
|---|---|---|---|
| Working Memory * | 1.665 + 0.197 | 1.122 + 0.317 | 1.515 + 0.157 |
| Vigilance ** | 0.226 + 0.089 | 0.251 + 0.093 |
p=0.000, 0.193 and 0.000 for Working Memory Task-1 vs 2, 1 vs 3 and 2 vs 3, respectively (Post-hoc Comparisons using Turkey’s HSD)
| Resting Sittin g | Auditory Task-1 | Auditory Task-2 | Working Memory Task-1 | Working Memory Task-2 | Working Memory Task - 3 | Vigilance Task-1 | Vigilance Task - 2 | ||
|---|---|---|---|---|---|---|---|---|---|
| RR Interva | 749 ± 88 | 762 ± 86 | 752 ± 84 | 752 ± 88 | 763 ± 82 | 748 ± 86 | 729 ± 92 | 741 ± 77 | |
| LF (0.04-0.15 Hz) | 1575 ± 774 | 1404 ± 881 | 1296 ± 1248 | 868 ± 485 | 848 ± 141 | 942 ± 536 | 1065 ± 835 | 1142 ± 592 | |
| MF (0.07-0.14 Hz) | 1005 ± 616 | 1035 ± 820 | 917 ± 1034 | 585 ± 395 | 555 ± 316 | 629 ± 404 | 764 ± 576 | 715 ± 424 | |
| HF (0.15-0.40 Hz) | 824 ± 791 | 826 ± 895 | 843 ± 1430 | 730 ± 662 | 669 ± 588 | 550 ± 380 | 611 ± 546 | 527 ± 421 | |
| Total Power (0.04-0.40 Hz) | 2339 ± 1361 | 2231 ± 1668 | 2139 ± 2658 | 1598 ± 964 | 1517 ± 923 | 1492 ± 816 | 1776 ± 1174 | 1669 ± 832 | |
| RR Interval | LF | MF | HF | Total Power | |
|---|---|---|---|---|---|
| Resting Sitting vs Auditory Task-1 | 0.379 | 0.609 | 0.955 | 0.910 | 0.532 |
| Resting Sitting vs Auditory Task-2 | 0.910 | 0.027 | 0.036 | 0.460 | 0.036 |
| Resting Sitting vs Working Memory Task-1 | 0.691 | 0.002 | 0.004 | 0.460 | 0.008 |
| Resting Sitting vs Working Memory Task-2 | 0.307 | 0.001 | 0.001 | 0.307 | 0.009 |
| Resting Sitting vs Working Memory Task-3 | 1.000 | 0.005 | 0.036 | 0.211 | 0.015 |
| Resting Sitting vs Vigilance Task-1 | 0.201 | 0.023 | 0.027 | 0.088 | 0.027 |
| Resting Sitting vs Vigilance Task-2 | 0.496 | 0.027 | 0.047 | 0.0487 | 0.017 |
| Auditory Task-1 vs 2 | 0.363 | 0.363 | 0.125 | 0.047 | 0.156 |
| Working Memory Task-1 vs 2 | 0.173 | 0.776 | 0.865 | 0.820 | 0.394 |
| Working Memory Task-1 vs 3 | 0.590 | 0.334 | 0.532 | 0.281 | 0.910 |
| Working Memory Task-2 vs 3 | 0.050 | 0.363 | 0.211 | 0.865 | 0.955 |
| Vigilance Task-1 vs 2 | 0.047 | 0.532 | 0.955 | 0.112 | 0.910 |
| Resting Sitting | Audit Task-1 | Audit Task-2 | Working Memory Task - 1 | Working Memory Task - 2 | Working Memory Task - 3 | Vigilance Task-1 | Vigilance Task-1 | |
|---|---|---|---|---|---|---|---|---|
| LF (0.04-0.15 Hz) | 71 ± 18 | 68 ± 15 | 69 ± 12 | 59 ± 17 | 60 ± 16 | 63 ± 13 | 67 ± 18 | 69 ± 17 |
| MF (0.07-0.14 Hz) | 38 ± 13 | 38 ± 11 | 37 ± 7 | 30 ± 9 | 31 ± 7 | 33 ± 10 | 35 ± 10 | 34 ± 9 |
| HF (0.15-0.40 Hz) | 29 ± 18 | 32 ± 15 | 31 ± 12 | 41 ± 17 | 40 ± 16 | 37 ± 13 | 33 ± 18 | 31 ± 17 |
| LF/HF Ratio | 4.32 ± 3.64 | 2.76 ± 1.66 | 2.77 ± 1.86 | 1.90 ± 1.24 | 1.95 ± 1.26 | 2.30 ± 1.90 | 3.06 ± 2.63 | 3.67 ± 3.48 |
| LF | MF | HF | LF/HF Ratio | |
|---|---|---|---|---|
| Resting Sitting vs Auditory Task-1 | 0.307 | 0.910 | 0.307 | 0.078 |
| Resting Sitting vs Auditory Task-2 | 0.394 | 0.910 | 0.394 | 0.069 |
| Resting Sitting vs Working Memory Task-1 | 0.011 | 0.041 | 0.011 | 0.008 |
| Resting Sitting vs Working Memory Task-2 | 0.023 | 0.041 | 0.023 | 0.031 |
| Resting Sitting vs Working Memory Task-3 | 0.125 | 0.394 | 0.125 | 0.047 |
| Resting Sitting vs Vigilance Task-1 | 9,140 | 0.691 | 0.140 | 0.125 |
| Resting Sitting vs Vigilance Task-2 | 0.609 | 0.691 | 0.609 | 0.650 |
| Auditory Task-1 vs 2 | 0.820 | 0.532 | 0.820 | 0.865 |
| Working Memory Task-1 vs 2 | 0.910 | 0.650 | 0.910 | 1.000 |
| Working Memory Task-1 vs 3 | 0.307 | 0.233 | 0.307 | 0.256 |
| Working Memory Task-2 vs 3 | 0.307 | 0.173 | 0.307 | 0.363 |
| Vigilance Task-1 vs 2 | 0.211 | 0.532 | 0.211 | 0.088 |
| Resting Sitting | Audit Task-1 | Audit Task-2 | Working Memory Task - 1 | Working Memory Task - 2 | Working Memory Task -3 | Vigilance Task-1 | Vigilance Task-1 | |
|---|---|---|---|---|---|---|---|---|
| LF (0.04-0.15 Hz) | 2.68 ± 1.10 | 2.39 ± 1.22 | 2.17 ± 1.73 | 1.53 ± 0.77 | 1.43 ± 0.63 | 1.65 ± 0.81 | 2.13 ± 1.20 | 2.02 ± 0.89 |
| MF (0.07-0.14 Hz) | 1.695 ± 0.85 | 1.72 ± 1.14 | 1.511 ± 1.43 | 1.013 ± 0.61 | 0.940 ± 0.49 | 1.083 ± 0.59 | 1.374 ± 0.78 | 1.260 ± 0.65 |
| HF (0.15-0.40 Hz) | 1.367 ± 1.18 | 1.351 ± 1.35 | 1.339 ± 120 | 1.238 ± 1.01 | 1.129 ± 0.97 | 0.951 ± 0.56 | 1.103 ± 0.97 | 0.922 ± 0.72 |
| LF/HF Ratio | 4.047 ± 1.93 | 3.743 ± 2.39 | 3.514 ± 3.67 | 2.769 ± 1.46 | 2.563 ± 1.45 | 2.603 ± 1.20 | 3.232 ± 1.75 | 2.941 ± 1.24 |
| LF | MF | HF | LF/HF Ratio | |
|---|---|---|---|---|
| Resting Sitting vs Auditory Task-1 | 0.532 | 0.955 | 0.910 | 0.496 |
| Resting Sitting vs Auditory Task-2 | 0.036 | 0.023 | 0.363 | 0.036 |
| Resting Sitting vs Working Memory Task-1 | 0.004 | 0.003 | 0.820 | 0.011 |
| Resting Sitting vs Working Memory Task-2 | 0.001 | 0.001 | 0.363 | 0.009 |
| Resting Sitting vs Working Memory Task-3 | 0.005 | 0.011 | 0.173 | 0.015 |
| Resting Sitting vs Vigilance Task-1 | 0.047 | 0.100 | 0.233 | 0.078 |
| Resting Sitting vs Vigilance Task-2 | 0.036 | 0.036 | 0.053 | 0.020 |
| Auditory Task-1 vs 2 | 0.363 | 0.125 | 0.156 | 0.078 |
| Working Memory Task-1 vs 2 | 0.609 | 0.865 | 0.496 | 0.394 |
| Working Memory Task-1 vs 3 | 0.307 | 0.427 | 0.307 | 0.865 |
| Working Memory Task-2 vs 3 | 0.334 | 0.156 | 1.000 | 0.733 |
| Vigilance Task-1 vs 2 | 0.570 | 0.650 | 0.047 | 0.334 |
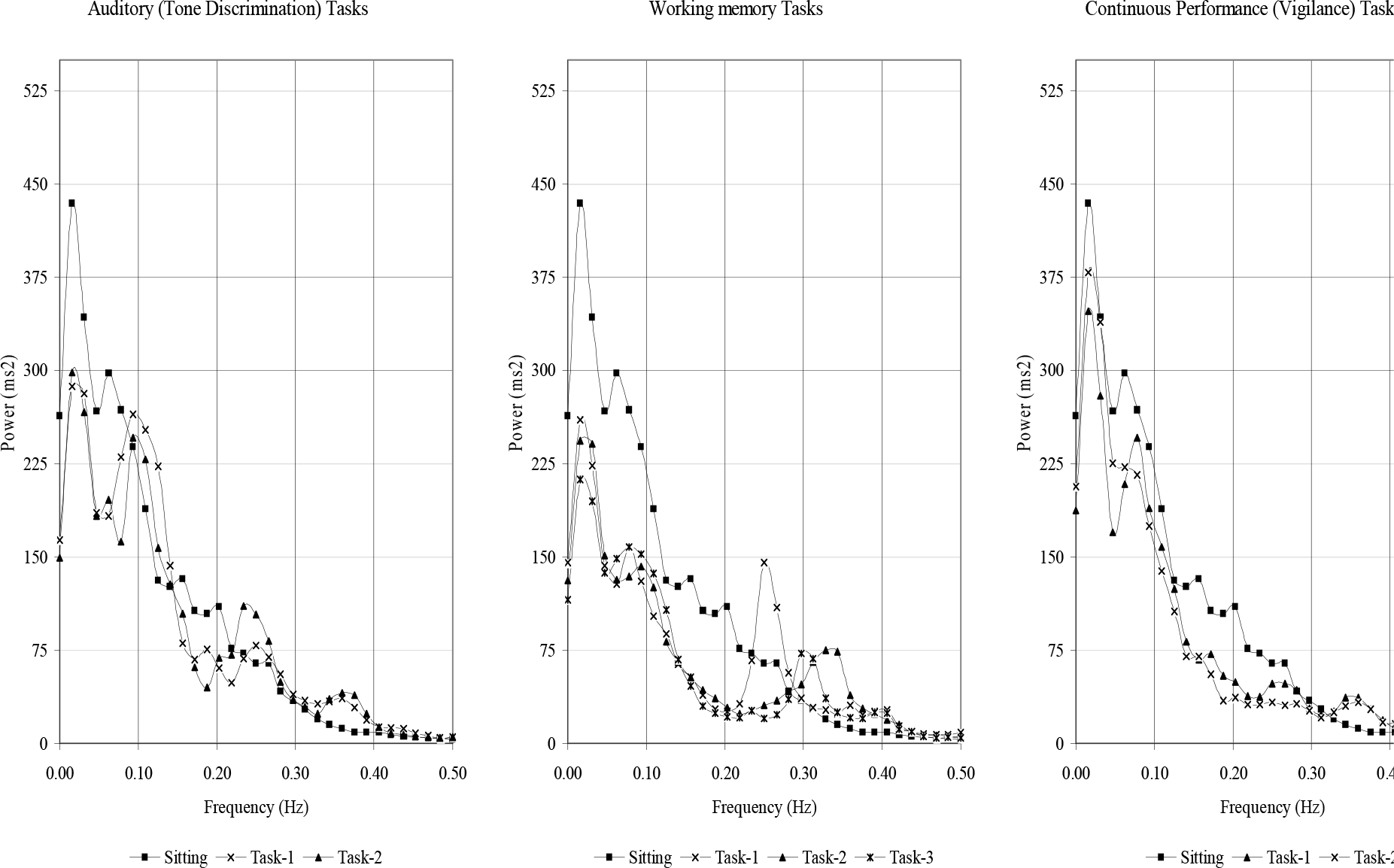
- Grand Average of HRV Spectra across the subjects (n=15)
Discussion
Auditory Tone discrimination Tasks-Time compression in Task-2 was manipulated to be 4 times more than that in Task-1 by decreasing the inter-stimulus delay from 2000 ms to 500 ms. Accuracy with which subjects could discriminate the two tones, was not different in the two conditions. Since increasing inter-stimulus interval is reported to increase the task difficulty and resources required to achieve a criterion level of performance [18], it can be reasonably construed that subjects invested more mental resources in Task-2 to attain comparable performance.
Working memory Tasks- Task difficulty/memory load in Task-2 was increased by decreasing the exposure time of target and array to 40% of the corresponding in Task-1, pre-target and inter-stimulus delay were not changed. The manipulation also amounts to an effective degradation of stimulus quality and absence of preview-the two variables reported to increase the task difficulty and complexity, respectively [18]. Target and array in these (working memory) tasks inflict concurrent (and mutually interfering) loads on working memory and, therefore, can be considered analogous to classical concurrent working memory load methods. Recall of target(s) is expected to be influenced by the viability of rehearsal in the early retention interval. A much reduced exposure time of stimuli in Task-2 permitted less time for such rehearsals and resulted into less efficient recall (accuracy was significantly less in Task-2). However, response time was significantly reduced in Task-2 compared to that in Task-1. In such instance, subjects are reported to respond as soon as possible to facilitate an effective recall [19]. In Task-3, time compression was increased by decreasing the pre-target and inter-stimulus delay to 40% of the corresponding valued in Task-1, Despite this reduction in pre target and inter-stimulus delay in Task-3, accuracy was preserved (was not different from that in Task-1, statistically) due to a comparable time span available for rehearsals because exposure time which could influence loads on to the working memory were identical. The apparent time compression (due to reduction in pre-target and inter-stimulus delay) was not appreciated by the subjects because of the unique feature of the tasks (vide, supra),
Vigilance Tasks - Compared to corresponding values in Task-1, both task difficulty and time compression were increased in Task-2 by decreasing the exposure time of target from 400 to 200 ms and inter-stimulus delay from 1000 to 500 ms. Accuracy and latency with which subjects could respond, were not different in the two conditions. It is reasonable to infer that subjects invested more mental resources in Task-2 to attain comparable performance (in this case, both accuracy and latency with which the subjects responded). In this task, response latency did not add to task length (vide supra).
HRV Indices - (Fig -1 and Tables 2 to 4 refer)
Expressed in absolute terms (ms2), total spectral power and power in LF and MF bands, exhibited significant rest-to-task differences (except in Auditory Task-2). No co-variation of these variables was demonstrable with further manipulations in cognitive demands between/amongst two/more variants of any task (Table-1 refers). Normalisation of these values relative to squared mean RR interval was not more informative because mean RR interval did not change across the task conditions (Tables-2 and 4 refer, vide infra, for further elaboration).
LF/HF Ratio and values of low-mid-and high-frequency power, when normalised to total spectral power, were significantly different in the Working Memory Tasks-1 and 2 compared to resting sitting. However, these variables failed to demonstrate a significant difference between resting sitting versus other task conditions [viz, Auditory (Tone Discrimination) Tasks and Vigilance Tasks] or between resting sitting and Working Memory Task-3 (differentiations between Resting Sitting and Working memory Task-3 by LF/HF ratio was just significant, p < 0.047). Normalised values of spectral power could also not discriminate between/ amongst two/more variants of the same task (Table-3 refers).
These observations, at the first instance, seem to institute superiority of expression of spectral values in absolute terms over that with normalisation for such applications as worked measurements. However, expression in absolute terms is not without the confounding influence of the changes in the total power on the values of different constituent components [11] and, thus, prevents appreciation of the fractional distribution of the spectral energy. [In the present study, power in high-frequency band, when expressed in absolute terms (as m sec2), did not differ significantly during Working Memory Task-1 and 2 compared to resting sitting. Nevertheless, its contribution to the total variability was identifiable to be significantly higher in the two conditions than in resting sitting when the corresponding values were normalised relative to total power].
It is noteworthy that mean RR interval did not change across different task conditions (Table-2 refers) and could not be accounted for significant rest-to-task differences observed in the study.
It has been argued that cardiac variability will decrease predominantly if rhythmic motor behaviour is required, for instance, during paced forced-choice discrimination tasks [20, 21]. Entrainment of stimulus frequency in the power spectrum of non linear physiologic signals is a well recognised phenomenon [22]. Experiments by Charnock and Manenica [23] further indicate existence of some psychophysiologic mechanism whereby subjects may adapt the pattern of their heart rate to externally imposed rhythms. In the present work, a paced rhythmic motor behaviour was elemental in the Working Memory Tasks wherein subjects were required to respond after a regular interval which was a sum of pre-target delay, exposure time of target, pre-array delay, exposure time of array, inter-stimulus delay and average reaction time. This interval was 4,650 ms, 3,209 ms and 3,972 ms, in Working Memory Tasks-1, 2 and 3, respectively. This corresponded to a frequency of 0.22, and 0.25 Hz, respectively. Grand averages of HRV spectra across the subjects reveal dominant components at these stimulus frequencies (Fig-1). The effect was remarkably absent in Auditory (Tone Discrimination) and Vigilance Tasks wherein subjects were required to respond at irregular intervals due to intrinsic task characteristics. Exact mechanism of such as entrainment is not known and is presumed to be through breathing. Respiratory pattern also is demonstrated to change with an increase in difficulty [24]. Subjects breathe slowly and deeply under mental load [25]. In the present study, no spectral analysis of breathing frequency was attempted. However, breathing is normally confined to 0.20-0.50 Hz and a slow and deep breathing pattern invariably results in to an increase in both HF-and LF-power (in absolute terms) [26, 27, 28]. The changes observed in the present study are in the an opposite direction, there was a significant reduction in total and LF-power observed in the present study is not attributable to changes in breathing pattern. The above noted dominance (or packing) of power at stimulus frequencies is also not due to significant increase in HF power. It is merely because of confinement of power in a narrower band.
A statistically significant difference observed in LF and MF components (expressed as values normalised relative to Total Power) of HRV between resting sitting and Working Memory Tasks-1 or 2 may seem, at the first instance, to be reciprocal to changes in HF component due to expression of spectral values in normalised units. However, significant rest-to-task differences in these components in HRV are also noticeable (Table-2 refers) even when expressed in absolute terms (m sec2) and not only when expressed in units normalised relative to Total Power. Furthermore, a statistical independence between RSA and slow wave alterations (0.02-0.07 Hz) in relation to mental load has been demonstrated [29]. Therefore, susceptibility of LF components to working memory loads can not be denied.
The results indicate an obvious sensitivity of the spectral components of HRV to working memory loads. However, a change was demonstrable only between ‘rest’ and ‘task’ conditions and it did not co-vary with a further manipulation of load in the working memory. These observations are in agreement with the findings of other studies conducted in controlled laboratory conditions [16, 30]. The observation of Jorna [16]- that load on working memory was one of the few task parameters to reveal HRV effects-is in conformity with the reported sensitivity of subjective appraisal of load in working memory [31]. Processing information in working memory is attention demanding and is referred to as controlled processing [32]. Tasks seem to be differentiated by HRV if they differ significantly in this respect.
Jorna [16], in an exhaustive review on spectral analysis of heart rate and psychological state, has confirmed an apparent lack of covariance of HRV with further manipulations of task characteristics despite a rather consistent rest-task effects. In his conception an increase in task demands is not accompanied by changes in HRV as long as the manipulation affects only structural or computational characteristics in the human information processing system. Changes in load will expectably be indexed by heart rate spectra parameters only if human limits are taxed as in dual-task settings [16]. This explains why a change in HRV components was not significant during Auditory (Tone Discrimination) and Vigilance Tasks. Jorna [16] also proposed that subjects select a particular psychological state which is perceived to be fit for the task and seem to be unwilling to change that.
Several mechanisms are believed to be involved in the regulations of HRV under mental load. The amplitude of the mid band is reduced because the HRV is less determined by changes in BP. The baroreceptor responses following decerebration is absent [33]. It is postulated [34] that decreased baroreceptor sensitivity (with a resultant effect on Traube-Herring-Meyer (THM) wave rhythmicity i.e. in 0.08-0.15 Hz band) reduces the inhibition of the Reticular Activating System (RAS) which, in turn increases the attention. In human subjects, a reduction of overall gain (around 0. 10 Hz band) of baroreceptor mechanisms has been demonstrated during mental arithmetic [35]. A significant effect of task difficulty on modulus in the mid band (0.07-0.14 Hz) has been reported during stimulated flying [25].
Results of the present study do establish susceptibility of certain spectral components (especially in the region 0.04-0.15 Hz) of HRV. The effects could not be explained on account of entrainment of power spectrum of HRV with the stimulus frequency and/or changes breathing pattern. Neither could a change in RR interval (with a consequent change in spectral power) account for this susceptibility. Nevertheless, attentional modulation of HRV seemed to be correlational rather than causational.
References
- Spectral analysis of sinus arrhythmia during mental loading. Ergonomics. 1975;18:255-70.
- [Google Scholar]
- Cardiac rhythmicities and attention in young children. Psychophysiology. 1997;34:547-52.
- [Google Scholar]
- Heart period variability of trained and untrained men at rest and during mental challenge. Psychophysiology. 1998;35:16-22.
- [Google Scholar]
- Heart Rate variability during attention phases in young infants. Psychophysiology. 1991;28:43-53.
- [Google Scholar]
- Heart rate variability : A new index of driver alertness/fatigue. In: Society of Automotive Engineers, New York. 1972. Report No 720141
- [Google Scholar]
- Air-to ground training missions; a psychophysiological workload analysis. Ergonomics. 1993;36:1071-87.
- [Google Scholar]
- Psychological responses to changes in workload during simulated air traffic control. Biological Psychology. 1996;37:361-77.
- [Google Scholar]
- A comparison of heart rate, eye activity, EEG and subjective measures of pilot mental workload during flight. Aviation Space and Environmental Medicine. 1998;69:360-7.
- [Google Scholar]
- Assessment of autonomic in humans by heart rate spectral analysis. Am J Physiol. 1985;248:11151-153.
- [Google Scholar]
- Power spectral analysis of heart rate and arterial pressure variabilities as a maker of sympatho-vagal interaction in man and conscious dog. Cir Res. 59:178-93.
- [Google Scholar]
- (1980). Power spectral analysis of heart rate variability in traumatic quadriplegic humans. American Journal of Physiology. 1980;258:111722-1726.
- [Google Scholar]
- Power spectrum analysis of heart rate fluctuations. A quantitative probe of beat-to-beat cardiovascular control. Science. 1981;213:220-22.
- [Google Scholar]
- Spectral analysis of heart rate and psychological state: A review of its validity as a workload index. Special Issue Cardiorespiratory measures and their role in studies of performance. Biological Psychology. 1992;34:237-57.
- [Google Scholar]
- Respiratory sinus arrhythmia in humans: an obligatory role for vagal feedback from the lungs. Journal of Applied Physiology. 1995;78:638-645.
- [Google Scholar]
- Attention In: Hancock PA, ed. Advances in Psychology - 47 - Human factors psychology. Amsterdam: Elsevier Science Publishers BV; 1987.
- [Google Scholar]
- Short Term Memory Limits in Human Performance In: Hancock PA, ed. Advances in Psychology - Vol 47 - Human factors psychology. Amsterdam: Elsevier Science Publishers BV; 1987.
- [Google Scholar]
- External and internal attention environments II, Reconsideration of the relationship between sinus arrhythmia and information load. Ergonomics. 1982;25:121-236.
- [Google Scholar]
- The use of entrainment in analysis of the human thermoregulatory system. J of Physiology. 1972;229:40-41.
- [Google Scholar]
- Spectral analysis of RR intervals under different work conditions. Ergonomics. 1978;21:103-8.
- [Google Scholar]
- Respiration in psychophysiology. Methods and applications. Biological Psychology. 1992;34:179-204.
- [Google Scholar]
- Physiological indices of workload in a simulated flight task. Biological Psychology. 1996;42:323-42.
- [Google Scholar]
- (1993). Important influence of respiration on human R-R interval power spectra is largely ignored. J Appl Physiol. ;75:2310-7.
- [Google Scholar]
- (1995). Override of spontaneous respiratory pattern generator reduces cardiovascular parasympathetic influence. J App Physiol. ;78:1048-1054.
- [Google Scholar]
- (1999). Effect of respiration rate on short term heart rate variability. J Clin Basic Cardiol. ;2:92-5.
- [Google Scholar]
- Respiratory and cardiac rhythms as windows to central and automatic bio-behavioural regulation: Selection of window frames, keeping the pane clean and viewing the neural topography. Biological Psychology. 1992;34:131-61.
- [Google Scholar]
- Metabolic and cardiorespiratory measures of mental effort: The effects of level of difficulty in a working memory task. International Journal of Psychophysiology. 1994;16:57-68.
- [Google Scholar]
- Subjective workload assessment and voluntary control of effort in a tracking task. Proceedings of the Annual Conference on Manual Control. 1984;20:57-72.
- [Google Scholar]
- Controlled and automatic human information processing II: Perceptual learning, automatic attending and a general theory. Psychological Review. 1977;84:127-90.
- [Google Scholar]
- Sinus Arrhythmia and Mental Workload In: Neville Moray, ed. Mental Workload, its theory and measurement. New York: Plenum press; 1979.
- [Google Scholar]
- Some autonomic-central nervous system interrelationships In: P Black, ed. Physiological correlates of emotion. New York: Academic Press;
- [Google Scholar]
- Assessment of the neutral control of the circulation during psychological stress. Journal of Autonomic Nervous System. 1991;35:33-41.
- [Google Scholar]




